
 Болезни
Болезни  Педиатрия
Педиатрия Первая помощь
Первая помощь Анатомия
Анатомия Питание
Питание Препараты
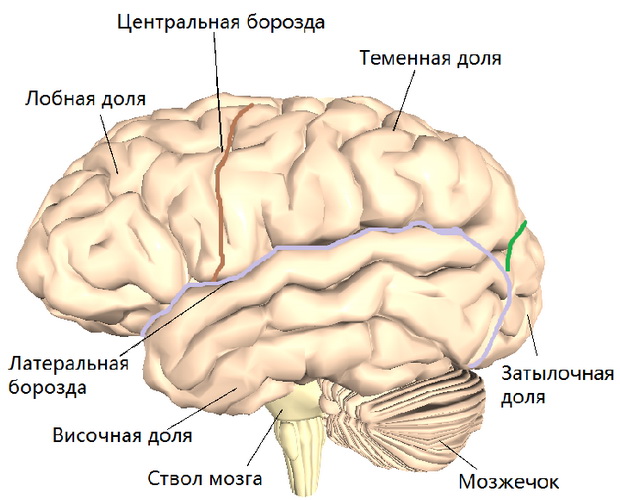
ПрепаратыОтдел головного мозга, находящийся в самой передней части черепной коробки, получил название конечного. Именно в этом отделе расположены такие жизненно важные центры, как центр высшей нервной деятельности и возникновения условных рефлексов, регуляции движений и произношения речи, центры зрения, слуха, обоняния, вкуса, а также кожной и мышечной чувствительности.
Конечный мозг продольной щелью делится на два полушария hemispheriae cerebrates, соединяющиеся друг с другом посредством системы спаек.
Каждое полушарие конечного мозга состоит из пяти долей:
- Лобную
- Височную
- Теменную
- Затылочную
- Островковую.
Полушария конечного мозга имеют сложный рельеф, обусловленный наличием борозд и извилин. Поверхность полушарий покрыта серым веществом — корой большого мозга.
К элементам внутреннего строения отделов конечного мозга относятся базальные ядра, nuclei basales, белое вещество, полостью каждого полушария является парный боковой желудочек, ventriculus lateralis.
В этой статье подробно описано анатомическое строение и основные функции конечного мозга.
Строение коры полушарий большого головного мозга человека: клетки и зоны
Кора полушарий большого мозга, cortex cerebri, составляет важнейшую часть органа ЦНС, являясь материальным субстратом высшей нервной деятельности и главным регулятором всех жизненных функций организма. Кора осуществляет анализ и синтез поступающих раздражений из внутренней среды организма и из окружающей внешней среды. Следовательно, с корой полушарий большого мозга связаны высшие формы отражения внешнего мира и сознательная деятельность человека.
В филогенетическом отношении выделяют древнюю (paleocortex), старую (archeocortex) и новую (neocortex) кору больших полушарий головного мозга. Древняя и старая кора располагаются на медиальной и базальной поверхности полушария. Их окружают промежуточные корковые формации, входящие в строение коры больших полушарий головного мозга и выделенные под названием перипалеокортекса и периархикортекса (mesocortex)
Самый наружный слой — молекулярная пластинка, lamina molecularis, содержит небольшое количество мелких нервных клеток и слагается преимущественно из густого сплетения нервных волокон, лежащих параллельно поверхности извилин.
Второй слой — наружная зернистая пластинка, lamina granulans externa, содержит большое количество мелких, полигональных или круглых нервных клеток.
Третий слой — наружная пирамидная пластинка, lamina pyramidalis externa, состоит из таких же мелких клеток, как и второй слой.
Четвертый слой называется внутренней зернистой пластинкой, lamina granularis interna.
Пятый слой — слой больших пирамидных клеток коры полушарий большого мозга или ганглиозный, laminaganglionaris, представлен внутренней пирамидной пластинкой, lamina pyramidalis interna. Он содержит наряду с довольно крупными пирамидными клетками, еще так называемые гигантские пирамидные клетки Беца, встречающиеся лишь в определенных участках коры: в передней центральной извилине (преимущественно в верхнем ее отделе) и в парацентральной I дольке медиальной поверхности полушария. Пирамидные клетки своей верхушкой обращены к поверхности мозга; основанием, от которого отходит аксон, — к белому веществу. Пятый слой дает начало эфферентным (нисходящим) корково-спинномозговому и корково-ядерному трактам.
Последний слой, лежащий на границе белого вещества, — полиморфный, lamina multiformis. В строение этого слоя коры полушарий большого мозга, как показывает его название, входят клеточные элементы самой разнообразной формы (треугольные, полигональные, овальные, веретенообразные).
Зону трех наружных слоев коры полушарий большого мозга принято объединять под названием главной наружной зоны. Три внутренних слоя коры больших полушарий головного мозга — это главная внутренняя зона.
Деятельность коры больших полушарий головного мозга: основные функции
Основные функции коры больших полушарий мозга определяются клеточным составом и межнейрональными связями пластинок. В молекулярной пластинке заканчиваются волокна из других слоев коры и из противоположного полушария. Существует мнение, что нейроны молекулярной пластинки имеют непосредственное отношение к процессам памяти. Наружная зернистая и наружная пирамидная пластинки в основном содержат ассоциативные нейроны, осуществляющие внутрикорковые связи. Они обеспечивают аналитические мыслительные процессы. Эти пластинки филогенетически наиболее молодые, они сильно развиты в коре полушарий большого мозга у человека. Внутренняя зернистая пластинка является главным афферентным слоем коры.
На нейронах этой пластинки заканчиваются проекционные нервные волокна, идущие от ядер таламуса и коленчатых тел. От пирамидных клеток внутренней пирамидной пластинки начинаются эфферентные проекционные волокна коры. Мультиформная пластинка содержит функционально неоднородные нейроны. От них берут начало ассоциативные и комиссуральные волокна.
Наряду с горизонтальной организацией коры в форме пластинок в настоящее время рассматривают принцип вертикальной модульной организации коры. В настоящее время получены данные о структурно-функциональной взаимосвязи клеток в различных слоях коры полушарий большого мозга. В связи с этим введено понятие корковых колонок, или модулей. Основу модулей составляют такие конструктивные заготовки, как колонки нейронов и пучки их апикальных дендритов.
Каждая корковая колонка представляет собой вертикально ориентированный ряд нейронов, проходящий через все слои коры. Принято считать, что в коре мозга существуют две разновидности стабильных генетически обусловленных объединений нейронов: микро- и макроколонки. В процессе жизнедеятельности из них могут формироваться функционально подвижные и варьирующие по структуре модули нейронов.
Микроколонки считаются основной модульной субъединицей в коре. Она представляет собой вертикально ориентированный тяж клеток, состоящий примерно из 110 нейронов и проходящий через все пластинки коры. Корковые колонки представляют собой модули, единицы обработки информации, обладающие собственным входом и выходом. Диаметр колонок составляет около 30 мкм. Почти во всех областях коры количество нейронов в колонках относительно постоянно, и только в корковых центрах зрения количество нейронов в колонках больше. Несколько сотен микроколонок объединяются в более крупную структуру — макроколонку, имеющую диаметр от 500 до 1000 мкм. Корковые колонки окружены радиально расположенными нервными волокнами и кровеносными сосудами.
Каждый такой модуль рассматривается как фокус конвергенции нескольких тысяч локальных, ассоциативных и каллозальных волокон. Между корковыми колонками и подкорковыми образованиями существуют топографически упорядоченные нервные связи, отдельным колонкам соответствуют определенные группы нейронов в базальных ядрах, таламусе, коленчатых телах.
Наиболее простыми и константными объединениями элементов нейронов являются пучки дендритов. Вертикальные пучки дендритов выполняют, по-видимому, основную конструктивную роль в консолидации нейронов. Деятельность коры больших полушарий головного мозга осуществляется, главным образом, аксонными терминалями релейных эфферентных волокон, а макроколонок — ассоциативными и каллозальными волокнами.
Отдельные дендриты в пучке могут на значительном протяжении непосредственно прилегать друг к другу, что создает благоприятные условия для реализации несинаптических влияний обмена ионами и метаболитами. В объединениях нейронов, образованных с помощью пучков дендритов, создаются структурные предпосылки как для дивергенции, так и для конвергенции синаптических импульсов.
С точки зрения миелоархитектоники в коре выделяют радиальные и тангенциальные нервные волокна. Первые вступают в кору из белого вещества, или наоборот, выходят из коры в белое вещество. Вторые располагаются параллельно поверхности коры и образуют на определенной глубине сплетения, называемые полосками. Различают полоски молекулярной пластинки, наружной и внутренней зернистых пластинок, внутренней пирамидной пластинки. Функции волокон коры больших полушарий головного мозга, проходящих в полосках, заключаются в связывании между собой нейронов соседних корковых колонок. Число полосок в различных полях коры неодинаково. В зависимости от него различают однополосковый, двухполосковый и многополосковый тип коры. Особенно хорошо выражены полоски в затылочной доле, в зрительных полях (стриарная кора).
На основании многочисленных клинических, патологоанатомических, электрофизиологических и морфологических исследований со всей определенностью установлено функциональное значение различных областей коры полушарий большого мозга.
Нервные центры коры больших полушарий головного мозга
Участки коры полушарий, имеющие характерную цитоархитектонику, и нервные связи, участвующие в выполнении определенных функций, являются нервными центрами. Поражение таких участков коры проявляется в утрате присущих им функций. Нервные центры плаща могут быть разделены на проекционные и ассоциативные.
Проекционные центры коры больших полушарий головного мозга — это участки, представляющие собой корковую часть анализатора, имеющие непосредственную морфофункциональную связь через посредство афферентных или эфферентных нервных путей с нейронами подкорковых центров.
Ассоциативные центры — это участки коры больших полушарий головного мозга человека, не имеющие непосредственной связи с подкорковыми образованиями, а связанные временной двусторонней связью с проекционными центрами. Ассоциативные центры играют первостепенную роль в осуществлении высшей нервной деятельности. В настоящее время достаточно точно выяснена динамическая локализация некоторых функций коры полушарий большого мозга. Участки коры полушарий большого мозга, не являющиеся проекционными или ассоциативными центрами, участвуют в выполнении межанализаторной интегративной деятельности головного мозга.
Корковые поля неравноценны в функциональном отношении и их можно разделить на первичные, вторичные и третичные.
Первичные поля представляют собой четко отграниченные участки, которые соответствуют центральным частям анализаторов. В эти поля проходит по специфическим проекционным афферентным путям основная масса сигналов от органов чувств. Первичные поля характеризуются сильным развитием внутренней зернистой пластинки. Первичные поля связаны с реле-ядрами таламуса и ядрами коленчатых тел. Они имеют экранную структуру и, как правило, жесткую соматотопическую проекцию, при которой отдельные участки периферии проецируются в соответствующие им участки коры. Повреждение первичных полей коры сопровождается нарушением непосредственного восприятия и тонкой дифференцировки раздражений.
Вторичные поля коры примыкают к первичным полям. Их можно рассматривать как периферические части корковых анализаторов. Эти поля связаны с ассоциативными ядрами таламуса. При поражении вторичных полей сохраняются элементарные ощущения, но нарушается способность к более сложным восприятиям. Вторичные поля не имеют четких границ, в них не выражена соматотопическая проекция.
Третичные поля коры отличаются наиболее тонкой нейронной структурой и преобладанием ассоциативных элементов. Эти поля связаны сзадними ядрами таламуса. В третичных полях осуществляются наиболее сложные взаимодействия анализаторов, лежащие в основе познавательного процесса (гнозия), формируются программы целенаправленных действий (праксия).
Кора обеспечивает совершенную организацию поведения животных на основе врожденных и приобретенных в онтогенезе функций и имеет следующие морфофункциональные особенности:
- Многослойность расположения нейронов;
- Модульный принцип организации;
- Соматотопическая локализация рецептирующих систем;
- Экранность, т.е. распределение внешней рецепции на плоскости нейронального поля коркового конца анализатора;
- Зависимость уровня активности от влияния подкорковых структур и ретикулярной формации;
- Наличие представительства всех функций нижележащих структур ЦНС;
- Цитоархитектоническое распределение на поля;
- Наличие в специфических проекционных сенсорных и моторной системах вторичных и третичных полей с ассоциативными функциями;
- Наличие специализированных ассоциативных областей;
- Динамическая локализация функций, выражающаяся в возможности компенсаций функций утраченных структур;
- Перекрытие в коре большого мозга зон соседних периферических рецептивных полей;
- Возможность длительного сохранения следов раздражения;
- Реципрокная функциональная взаимосвязь возбудительных и тормозных состояний;
- Способность к иррадиации (распространению) возбуждения и торможения;
- Наличие специфической электрической активности.
Что включают в себя центральный и периферический отделы обонятельного мозга?
Обонятельный мозг – это сенсорная зона коры головного мозга, он развивается из вентральной части конечного мозга. Обонятельный мозг состоит из двух отделов: периферического и центрального.
Обонятельная доля или периферический отдел обонятельного мозга включает образования, расположенные на основании мозга:
- Обонятельную луковицу, bulbus olfactorius;
- Обонятельный тракт, tractus olfactorius;
- Обонятельный треугольник, trigonum olfactorium;
- Переднее продырявленное вещество, substantia perforata anterior.
Центральный отдел обонятельного мозга включает:
- Сводчатую извилину, gyrusfornica tus, которая заканчивается вблизи височного полюса крючком, uncus’,
- Ногу морского коня, или Аммонов рог, hippocampus, (cornu Ammoni) — особой формы образование, расположенное в полости нижнего рога бокового желудочка;
- Зубчатую извилину, gyrus dentatus, обнаруживаемую в виде узкой полоски в глубине борозды гиппокампа, под ногой морского коня.
В следующем разделе статьи описывается лимбическая система мозга, ее структура и функции.
Анатомическая структура лимбической системы головного мозга
Лимбическая система мозга представляет собой функциональное объединение структур мозга, участвующих в организации эмоционально-мотивационного поведения, таких как пищевой, половой, оборонительный инстинкты. Эта система участвует в организации цикла бодрствование-сон.
Латинское слово limbus означает кайма, край. Лимбическая система мозга человека названа так потому, что корковые структуры, входящие в нее, находятся на краю неокортекса и как бы окаймляют ствол мозга.
Лимбическая система головного мозга как филогенетически древнее образование оказывает регулирующее влияние на кору большого мозга и подкорковые структуры, устанавливая необходимое соответствие уровней их активности.
Так, лимбическая система имеет отношение к регулированию уровня реакции автономной, соматической систем при эмоционально-мотивационной деятельности, регулированию уровня внимания, восприятия, воспроизведения эмоционально значимой информации. Лимбическая система определяет выбор и реализацию адаптационных форм поведения, динамику врожденных форм поведения, поддержание гомеостаза, генеративных процессов. Наконец, она обеспечивает создание эмоционального фона, формирование и реализацию процессов высшей нервной деятельности.
В настоящее время хорошо известны связи между структурами лимбической системы мозга, организующие круги и имеющие свою функциональную специфику. К ним относится круг Пейпеса (гиппокамп — сосцевидные тела — передние ядра таламуса — кора поясной извилины — парагиппокампальная извилина — гиппокамп). Этот круг имеет отношение к памяти и процессам обучения.
Другой круг (миндалевидное тело — гипоталамус — мезенцефальные структуры — миндалевидное тело) регулирует агрессивно-оборонительные, пищевые и сексуальные формы поведения.
Считается, что образная (иконическая) память формируется кортико-лимбико-таламо-кортикальным кругом. Круги разного функционального назначения связывают лимбическую систему со многими структурами центральной нервной системы, что позволяет последней реализовать функции, специфика которых определяется включенной дополнительной структурой.
Например, включение хвостатого ядра в один из кругов лимбической системы определяет ее участие в организации тормозных процессов высшей нервной деятельности. Свои нисходящие пути лимбическая система направляет к ретикулярной формации ствола мозга и к гипоталамусу. Через гипоталамо-гипофизарную систему лимбическая система осуществляет контроль над гуморальной системой. Для лимбической системы характерна особая чувствительность и особая роль в ее функционировании гормонов, синтезируемых в гипоталамусе и секретируемых гипофизом, — окситоцина и вазопрессина.
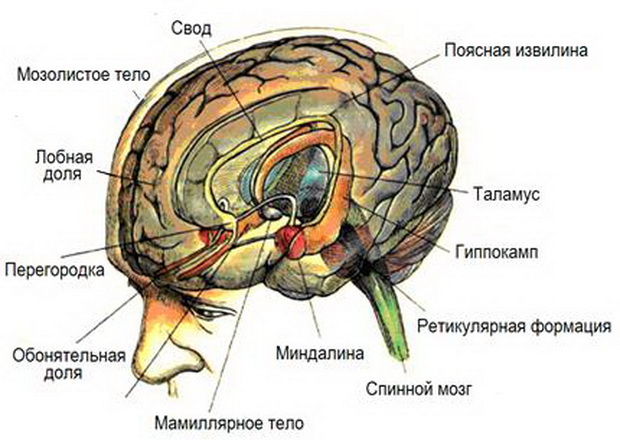
Какие функции выполняет лимбическая система в мозге человека?
Наиболее полифункциональными образованиями лимбической системы являются гиппокамп и миндалевидные тела. Физиология этих структур наиболее изучена.
Миндалевидное тело (corpus amigdaloideum) — подкорковая структура лимбической системы, расположенная в глубине височной доли мозга. Нейроны миндалины лимбической системы мозга разнообразны по функциям, форме и нейрохимическим процессам в них. Функции миндалевидного тела лимбической системы мозга связаны с обеспечением оборонительного поведения, вегетативными, двигательными, эмоциональными реакциями, мотивацией условнорефлекторного поведения.
Миндалевидное тело реагирует многими своими ядрами на зрительные, слуховые, интероцептивные, обонятельные, кожные раздражения, причем все эти раздражения вызывают изменение активности любого из ядер, т.е. ядра миндалевидного тела полисенсорны.
Гиппокамп (hippocampus), расположен в глубине височных долей мозга и является основной структурой лимбической системы. Он имеет своеобразную изогнутую форму (гиппокамп в переводе — морской конек) и почти на всем своем протяжении образует впячивание в полость нижнего рога бокового желудочка. Гиппокамп является собственно складкой (извилиной) старой коры. С ней сращена и заворачивается над ней зубчатая извилина. Многочисленные связи гиппокампа со структурами как лимбической системы, так и других отделов мозга определяют его многофункциональность, не подвергается сомнению его участие в ориентировочном рефлексе, реакциях настороженности, повышении внимания, в динамике обучения, что чаще наблюдается при высоком уровне эмоционального напряжения — страхе, агрессии, голоде, жажде.
Гипоталамус (hypothalamus) как структура промежуточного мозга, входящая в лимбическую систему мозга человека выполняет следующие функцию: организует эмоциональные, поведенческие, гомеостатические реакции организма. Гипоталамус имеет большое число нервных связей с корой большого мозга, подкорковыми узлами, зрительным бугром, средним мозгом, мостом, продолговатым и спинным мозгом. Организация афферентных и эфферентных связей гипоталамуса свидетельствует о том, что он служит важным интегративным центром для соматических, вегетативных и эндокринных функций.
Латеральные ядра гипоталамуса образуют двусторонние связи с верхними отделами ствола мозга, центральным серым веществом среднего мозга (лимбической областью среднего мозга) и с лимбической системой. Чувствительные сигналы от поверхности тела и внутренних органов поступают в гипоталамус по восходящим спинно-бульбо-ретикулярным путям.
Медиальные ядра гипоталамуса обладают двусторонними связями с латеральными и, кроме того, непосредственно получают ряд сигналов из остальных отделов головного мозга. В медиальной области гипоталамуса существуют особые нейроны, воспринимающие важные параметры крови и спинномозговой жидкости; иными словами, эти нейроны следят за состоянием внутренней среды организма. Они могут воспринимать, например, температуру крови («тепловые» нейроны), солевой состав плазмы (осморецепторы) или содержание гормонов в крови. Посредством нервных механизмов медиальная область гипоталамуса управляет деятельностью нейрогипофиза, а посредством гормональных — аденогипофиза. Таким образом, эта область служит промежуточным звеном между нервной и эндокринной системами, представляя собой «нейроэндокринный интерфейс».
Подкорковые базальные ядра головного мозга
Базальные подкорковые ядра мозга, nuclei basales, представляют собой скопления серого вещества в нижних отделах полушарий. Они являются филогенетически старыми образованиями. Их выделяют в качестве стволовой части конечного мозга. К базальным ядрам относят полосатое тело, ограду и миндалевидное тело.
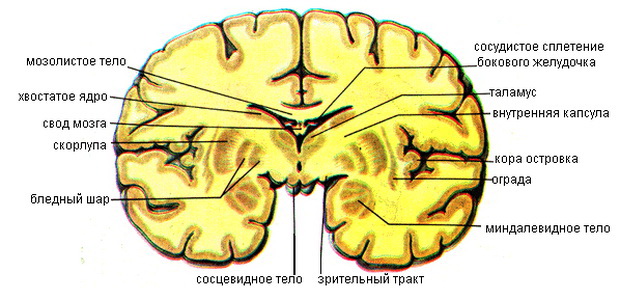
Из подкорковых ядер конечного головного мозга хвостатое ядро и скорлупа объединяются под названием полосатое тело, corpus striatum, а вместе с бледным шаром, globus pallidus, они составляют так называемую стриопаллидарную систему. Такое объединение обусловлено функциональной взаимосвязью. Указанные структуры взаимно уравновешивают друг друга и благодаря этому оказывают оптимальное влияние на двигательные акты.
Являясь высшим отделом экстрапирамидной системы, они обеспечивают выполнение различных непроизвольных (автоматизированных) движений, регулируют состояние мышечного тонуса, а, следовательно, влияют и на характер произвольных движений. Причем в единой функциональной системе паллидум оказывает активизирующее воздействие на подкорковые образования экстрапирамидной системы, а стриатум — тормозящее. Стриопаллидарная система получает афферентную информацию от нейронов медиального ядра таламуса.
Кроме того, стриарная система имеет связи с корой полушарий большого мозга, в частности, с корой лобной, височной и затылочной долей. В полосатом теле заканчивается эфферентный корково-стриарный путь, tractus corticostriatus. В свою очередь, стриарная система посылает тормозные эфферентные импульсы на нейроны бледного шара. От последнего эфферентные импульсы достигают нейронов двигательных ядер спинного мозга и черепных нервов. Следует отметить, что большая часть нервных волокон по пути следования от подкорковых узлов до клеток двигательных ядер переходит на противоположную сторону. Таким образом, подкорковые узлы каждого полушария большого мозга оказываются связанными в основном с противоположной половиной тела.
Стриопаллидарная система получает афферентные волокна от неспецифических медиальных таламических ядер, лобных отделов коры больших полушарий, коры мозжечка и черной субстанции среднего мозга. Основная масса эфферентных волокон стриатума радиальными пучками сходится к бледному шару. Таким образом, бледный шар является выходной структурой стриопаллидарной системы. Эфферентные волокна бледного шара идут к передним ядрам таламуса, которые связаны с фронтальной и теменной корой больших полушарий. Часть эфферентных волокон, не переключающихся в ядре бледного шара, идет к черной субстанции и красному ядру среднего мозга. Стриопаллидум, совместно со своими проводящими путями, входит в экстрапирамидную систему, оказывающую тоническое влияние на моторную деятельность. Эта система контроля над движениями называется экстрапирамидной потому, что переключается на пути к спинному мозгу, минуя пирамиды продолговатого мозга.
Стриопаллидарная система является высшим центром непроизвольных и автоматизированных движений, снижает мышечный тонус, тормозит движения, осуществляемые двигательной корой.
Базальные ядра головного мозга (правого и левого полушарий) связаны между собой комиссуральными волокнами, которые проходят в составе задней спайки мозга. Это обеспечивает их сочетанную работу по выполнению автоматизированных, обычно стереотипных, но довольно сложных рефлекторных двигательных актов, в том числе локомоторных (ходьба, плавание, прием пищи и т.д.), которые человек совершает «не думая».
Тесная связь стриопаллидарной системы с ядрами гипоталамуса (задняя группа ядер гипоталамуса) объясняет возможность ее влияния на эмоциональные реакции.
Хвостатое ядро (nucleus caudatus) и скорлупа (putamen) являются эволюционно более поздними (neostriatum), чем бледный шар (paleostriatum), образованиями и функционально оказывают на него тормозящее влияние. Обилие и характер связей хвостатого ядра и скорлупы свидетельствуют об их участии в интегративных процессах, организации и регуляции движений, регуляции работы вегетативных органов. Во взаимодействиях хвостатого ядра и бледного шара превалируют тормозные влияния. Взаимодействие черного вещества и хвостатого ядра основано на прямых и обратных связях между ними.
Бледный шар (globuspallidus, pallidum) имеет преимущественно крупные нейроны Гольджи I типа. Связи бледного шара с таламусом, скорлупой, хвостатым ядром, средним мозгом, гипоталамусом, соматосенсорной системой и др. свидетельствуют об его участии в организации простых и сложных форм поведения.
Ограда (claustrum) содержит полиморфные нейроны разных типов. Она образует связи преимущественно с корой большого мозга.
Глубокая локализация и малые размеры ограды представляют определенные трудности для ее физиологического исследования. Это подкорковое ядро головного мозга имеет форму узкой полоски серого вещества, расположенного под корой большого мозга в глубине белого вещества.
Известно, что толщина ограды левого полушария у человека несколько больше, чем правого; при повреждении ограды правого полушария наблюдаются расстройства речи.
Таким образом, базальные ядра головного мозга являются интегративными центрами организации моторики, эмоций, высшей нервной деятельности.
Миндалевидное тело является комплексом базальных ядер полушарий мозга, расположенных в переднем полюсе височной доли полушария и непосредственно соприкасающихся с корой парагиппокампальной извилины. К ним подходят волокна из обонятельного тракта, таламуса и коры. Эфферентные пути миндалевидного тела идут в терминальной полоске. Миндалевидное тело относится к лимбической системе.
Функции волокон белого вещества больших полушарий головного мозга
Волокна белого вещества полушарий мозга можно подразделить на три группы: ассоциативные, комиссуральные и проекционные.
Комиссуральные волокна соединяют симметричные части полушарий большого мозга. В отличие от ассоциативных они имеют преимущественно поперечный ход.
Ассоциативные волокна белого вещества мозга соединяют различные отделы коры в пределах одного полушария. Ассоциативные волокна, которые не выходят за пределы коры, называются интракортикальными ассоциативными волокнами. Те ассоциативные волокна, которые, соединяя отдельные участки коры, выходят из коры в белое вещество полушарий головного мозга, чтобы возвратиться в кору в другом месте, называются экстракортикальными ассоциативными волокнами. Они делятся на две группы — короткие и длинные.
Проекционные волокна связывают кору большого головного мозга с нижележащими отделами, пронизывая полушария в вертикальном направлении. Большинство проекционных путей проходит через внутреннюю капсулу.
К длинным ассоциативным путям относят:
- Верхний продольный пучок (fasciculus longitudinalis superior) — находится в верхней части белого вещества полушария большого мозга и соединяет кору лобной доли с теменной и затылочной.
- Нижний продольный пучок (fasciculus longitudinalis inferior) — находится в нижних отделах полушария и соединяет кору височной доли с затылочной.
- Крючковидный пучок (fasciculus uncinatus) — дугообразно изгибаясь впереди островка, соединяет кору лобного полюса с передней частью височной доли.
- Пояс (cingulum) — охватывает в виде кольца мозолистое тело и соединяет участки коры в лобной, затылочной и височной долях.
- Подмозолистый пучок (fasciculus subcallosus) — располагается кнаружи от поясного пучка и соединяет участки коры в лобных извилинах и в извилинах латеральной поверхности затылочной доли.
Проекционные нервные волокна, идущие от полушария большого мозга к нижележащим его отделам, составляют внутреннюю капсулу и ее лучистый венец (corona radiata). Книзу волокна нисходящих проводящих путей внутренней капсулы в виде компактных пучков направляются в ножку среднего мозга.
Через переднюю ножку внутренней капсулы проходят, во-первых, волокна белого вещества полушарий большого мозга, которые соединяют таламус с корой лобной доли. Это таламокортикальные и кортикоталамические пучки. Кроме того, через переднюю ножку внутренней капсулы проходят лобно-мостовые пути. Через колено внутренней капсулы проходят корково-ядерные волокна, то есть та часть двигательного пирамидного пути, которая проводит произвольные импульсы к сокращениям мышц головы и шеи.
В задней ножке внутренней капсулы рядом с коленом проходят волокна корково-спинномозгового (пирамидного) пути — той части главного двигательного пирамидного пути, которая проводит произвольные импульсы ксокращениям мышц из двигательного центра коры к передним рогам серого вещества спинного мозга. Рядом с корково-спинномозговым трактом в следующем отрезке задней части внутренней капсулы идут восходящие таламокортикальные волокна, которые, возникая в таламусе, направляются в теменную долю полушария. По ним проводятся чувствительные импульсы общего кожного и мышечного чувства. Еще дальше кзади в задней ножке внутренней капсулы проходят волокна височно-затылочно-мостового пути.
В той части внутренней капсулы, которая находится кзади от чечевицеобразного ядра, проходят волокна белого вещества больших полушарий головного мозга, возникающие в латеральном коленчатом теле и направляющиеся в зрительный центр коры. Наконец, в той части внутренней капсулы, которая расположена ниже чечевицеобразного ядра, проходят волокна слухового пути. Они начинаются в медиальном коленчатом теле и заканчиваются в корковом слуховом центре.
Таким образом, внутренняя капсула — это такой слой белого вещества большого мозга, который является действительно воротами для всех центростремительных и центробежных проекционных путей, которые идут к коре или от нее. Меньшее значение имеют наружная и крайняя капсулы. Здесь проходят главным образом ассоциативные пучки волокон.
Мозолистое тело (corpus callosum) содержит комиссуральные волокна, соединяющие кору правого и левого полушарий головного мозга. Верхняя поверхность мозолистого тела имеет серый покров, indusium griseum, и продольные полоски, striae longitudinales corporis callosi, входящие в состав обонятельного мозга. В мозолистом теле располагаются волокна, соединяющие новые, более молодые, отделы коры (neopaleum), корковые центры правого и левого полушарий, в которых волокна мозолистого тела расходятся веерообразно, образуя лучистость мозолистого тела (radiatio corporis callosi).
Функции комиссуральных волокон белого вещества головного мозга, идущих в колене и клюве мозолистого тела, заключаются в соединении друг с другом участков коры лобных долей правого и левого полушарий. Загибаясь кпереди, пучки этих волокон как бы охватывают с двух сторон переднюю часть продольной щели большого мозга и образуют лобные (большие) щипцы (forceps frontalis major). В стволе мозолистого тела проходят нервные волокна, соединяющие кору центральных извилин, теменных и височных долей двух полушарий мозга. Валик мозолистого тела состоит из комиссуральных волокон белого вещества мозга, его функции – соединение коры затылочных и задние отделы теменных долей правого и левого полушарий. Изгибаясь кзади, пучки этих волокон охватывают задние отделы продольной щели большого мозга и образуют затылочные (малые) щипцы (forceps occipitalis minor).
Под мозолистым телом находится свод, fornix, состоящий из двух тяжей: начинающихся от бахромки гиппокампа (fimbriae hippocampi) ножек, crus fornicis, соединенных в средней части спайкой свода, comissura fornicis, после чего формируется тело свода, corpus fornicis, которое расходится кпереди и книзу на два столба свода, columnae fornicis. Столбы свода заканчиваются в сосцевидных телах. Таким образом, свод мозга (эфферентный проекционный путь) соединяет кору височной доли (гиппокамп) с промежуточным мозгом (с сосцевидными телами гипоталамуса).
Боковые желудочки, левый (первый) и правый (второй), сообщаются с III желудочком посредством межжелудочкового отверстия, foramen interventriculare (Monroi). Через это отверстие из полости III желудочка в каждый боковой желудочек проникает сосудистое сплетение, plexus choroideus ventriculi lateralis, которое простирается в центральную часть, полость заднего и нижнего рогов. Со стороны желудочков сосудистое сплетение покрыто тонкой пластинкой эпендимы, выстилающей также стенки всех полостей. Сосудистые сплетения желудочков мозга продуцируют спинномозговую жидкость.
