
 Болезни
Болезни  Педиатрия
Педиатрия Первая помощь
Первая помощь Анатомия
Анатомия Питание
Питание Препараты
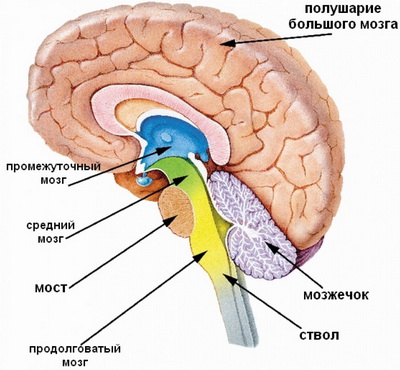
ПрепаратыВ макроанатомии ствола головного мозга (runcus encephali) выделяют 17 отделов: от подушки таламуса до задвижки на заднем поле. С анатомической точки зрения в строение этого протяжённого образования включают продолговатый, ромбовидный и средний мозг, стволовой (или варолиев) мост, а также мозжечок. Структурные особенности и основные функции каждого из этих отделов подробно рассмотрены на этой странице.
Ствол головного мозга человека — это филогенетически древняя часть, в которой располагаются структуры, относящиеся к сегментарному аппарату головного мозга, подкорковые центры слуха, зрения, обоняния и тактильной чувствительности. В структуру ствола головного мозга входят: продолговатый мозг, мост и средний мозг. С ними анатомически и функционально связаны 10 пар черепных нервов ствола головного мозга — III—XII. II пара черепных нервов — зрительный нерв, связана с промежуточным мозгом. I пара черепных нервов — обонятельные нервы — с конечным.
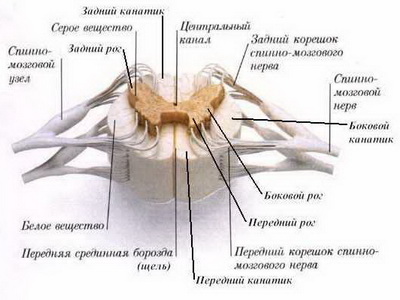
Сегментарный аппарат ствола головного мозга
Сегментарный аппарат ствола головного мозга представляет собой совокупность анатомически и функционально взаимосвязанных структур, предназначенных для осуществления безусловных (врожденных) рефлексов, замыкающихся на уровне ствола головного мозга. Примерами таких рефлексов являются сосательный, глотательный, роговичный, кашлевой и т.д.
В состав этого отдела ствола головного мозга входят следующие структуры:
- Корешковые волокна черепных нервов, включающих чувствительный компонент — V пара (тройничный нерв), VII пара (лицевой нерв), IX пара (языкоглоточный нерв), X пара (блуждающий нерв). Они представляют собой расположенные в веществе ствола головного мозга центральные отростки псевдоуниполярных клеток тройничного узла (V пары), узла коленца (VII пары), верхнего и нижнего узлов (IX и X пар). Корешковые волокна заканчиваются синаптическими окончаниями на вставочных нейронах ствола головного мозга.
- Вставочные нейроны, роль которых выполняют рассеянные клетки ретикулярной формации ствола головного мозга. Аксоны этих клеток синаптически заканчиваются на нейронах двигательных ядер черепных нервов.
- Мультиполярные нейроны двигательных ядер черепных нервов ствола головного мозга — III пара (глазодвигательный нерв), IV пара (блоковый нерв), V пара (тройничный нерв), VT пара (отводящий нерв), VII пара (лицевой нерв), IX пара (языкоглоточный нерв), X пара (блуждающий нерв), XI пара (добавочный нерв) и XII пара (подъязычный нерв).
- Часть аксонов нейронов двигательных ядер черепных нервов, составляющих двигательные корешковые волокна в пределах вещества мозга.
- Остальные элементы рефлекторных дуг безусловных рефлексов относятся к периферической нервной системе (корешковые волокна, лежащие за пределами ствола головного мозга, краниальные чувствительные ганглии, черепные нервы и их ветви).
В большинстве случаев вставочные нейроны сегментарного аппарата ствола головного мозга обеспечивают передачу нервных импульсов на нейроны двигательных ядер нескольких черепных нервов, причем не только своей стороны, но и противоположной. Например, при раздражении кожи лица в области щеки или губ у новорожденного возникают сосательные движения. Воспринимают раздражение рецепторы, являющиеся окончаниями псевдоуниполярных клеток узла тройничного нерва. Распространение нервного импульса в стволе головного мозга осуществляется на нейроны двигательных ядер V, VII, IХ, Х,ХI и ХII пар черепных нервов. В связи с этим в осуществлении такой функции ствола головного мозга, как сосательный акт, принимают участие жевательные, мимические мышцы, мышцы неба, глотки, шеи и языка. При этом мускулатура включается в осуществление ответной реакции в равной степени как на своей, так и на противоположной стороне тела.
Далее вы узнаете о строении и функциях продолговатого мозга.
Строение и основные функции продолговатого мозга
Продолговатый мозг, medulla oblongata (myelencephalon), находится между задним мозгом и спинным мозгом.
Верхняя граница продолговатого мозга на вентральной поверхности головного мозга проходит по нижнему краю моста, на дорсальной поверхности соответствует мозговым полоскам IV желудочка, которые делят дно желудочка на верхнюю и нижнюю части. Граница между продолговатым мозгом и спинным мозгом соответствует уровню большого затылочного отверстия или месту выхода из мозга верхней части корешков первой пары спинномозговых нервов.
При переходе от спинного мозга к продолговатому центральный канал расширяется в желудочек, которому на задней поверхности продолговатого мозга и моста соответствует ромбовидная ямка. Вследствие этого части, лежащие кзади от этого центра продолговатого мозга, смещаются в стороны. Серое вещество, располагающееся в спинном мозге в виде сплошных столбов, теряет свою непрерывность и распадается на отдельные ядра. Появляются новые ядра и связанные с ними системы нервных волокон.
На задней поверхности головного продолговатого мозга хорошо различимы продолжающиеся из заднего канатика спинного мозга тонкий и клиновидный пучки. Они оканчиваются в собственных ядрах, лежащих по обе стороны ромбовидной ямки. На латеральную поверхность продолговатого мозга продолжается боковой канатик с расположенными в нем спинно-мозжечковыми путями. В дорсальной части продолговатого мозга находится ретикулярная формация. Здесь же заложены ядра черепных нервов продолговатого мозга, с IX по XII пару. Большинство ядер проецируется на определенные участки ромбовидной ямки.
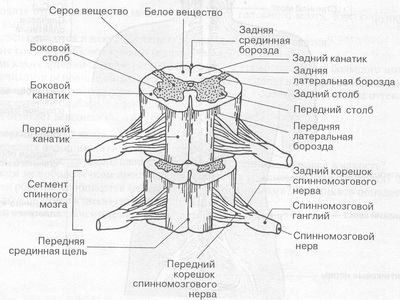 В строение передней поверхности продолговатого мозга продолговатого мозга находятся пирамиды и оливы. Пирамиды содержат корково-спинномозговые пути, на границе со спинным мозгом они совершают неполный перекрест, после которого перекрещенные волокна уходят в боковые канатики спинного мозга.
В строение передней поверхности продолговатого мозга продолговатого мозга находятся пирамиды и оливы. Пирамиды содержат корково-спинномозговые пути, на границе со спинным мозгом они совершают неполный перекрест, после которого перекрещенные волокна уходят в боковые канатики спинного мозга.
В оливах заложено несколько ядер (нижнее оливное, медиальное добавочное, заднее добавочное). Они получают эфферентные волокна из полосатого тела, ретикулярной формации. От ядер оливы берут начало оливо-мозжечковый и оливо-спинно-мозговой пути. Олива в отделах продлоговатого мозга осуществляет кольцевую обратную связь между стволом мозга и мозжечком и оказывает влияние на двигательные нейроны передних рогов спинного мозга.
От тонкого и клиновидного ядер начинаются внутренние дугообразные волокна, которые перекрещиваются в глубине продолговатого мозга и образуют восходящий путь, называемый медиальной петлей. К медиальной петле присоединяется спинно-таламический путь, в результате чего образуется система волокон, проводящих кожную и проприоцептивную Чувствительность к ядрам таламуса.
Задний спинно-мозжечковый путь в продолговатом мозге человека вступает в нижние мозжечковые ножки, а передний путь проходит через продолговатый мозг и мост до среднего мозга.
Перекрещенные волокна корково-спинномозгового пути уходят в боковые канатики спинного мозга, образуя латеральные корково-спинномозговые пути, а неперекрещенные волокна продолжаются в передние канатики в виде передних корково-спинномозговых путей.
Основные функции продолговатого мозга – участие за счет ядерных образований и ретикулярной формации в реализации вегетативных, соматических, вкусовых, слуховых, вестибулярных рефлексов. Особенностью продолговатого мозга является то, что его ядра, возбуждаясь последовательно, обеспечивают выполнение сложных рефлексов, требующих последовательного включения разных мышечных групп, что наблюдается, например, при глотании.
Такие образования головного мозга, как мост, средний мозг, мозжечок, таламус, гипоталамус и кора большого мозга, имеют двусторонние связи с продолговатым мозгом. Наличие этих связей свидетельствует об участии продолговатого отдела головного мозга в регуляции тонуса скелетной мускулатуры, вегетативных и высших интегративных функций, анализе сенсорных раздражений.
Мост головного мозга: строение и основные функции отдела
Задний мозг, metencephalon, включает мост, расположенный спереди (вентрально) и мозжечок, который находится позади моста. Полостью заднего мозга, а вместе с ним и продолговатого, является IV желудочек.
Мост как отдел головного мозга граничит сзади с продолговатым мозгом, а спереди — с ножками мозга.
Здесь заложены ядра черепных нервов, с V по VIII пары. В дорсальной части моста находится ретикулярная формация. Она является продолжением ретикулярной формации продолговатого мозга и началом этой же системы среднего мозга. Аксоны нейронов ретикулярной формации моста идут в мозжечок, в спинной мозг (ретикулоспинальный путь). Последние активируют нейроны спинного мозга. В передней части моста располагаются собственные ядра моста. К ним подходят корково-мостовые волокна из всех долей одноименного полушария большого мозга. От ядер моста отходят мосто-мозжечковые волокна, образуя среднюю мозжечковую ножку. Таким образом, основная функция моста головного мозга – это передача импульсов из коры большого мозга в кору мозжечка. Эта связь необходима для взаимодействия большого мозга и мозжечка в регуляции двигательных функций.
В строение моста головного мозга на вентральной поверхности входят:
- Базилярная борозда, sulcus basilaris, — в ней проходит одноименная артерия;
- Горизонтальная борозда, sulcus horizontalis, — является границей между мостом и продолговатым мозгом;
- Средние ножки мозжечка, pedunculi cerebellares medii.
Образования на дорсальной поверхности:
- Мост образует верхний угол ромбовидной ямки, fossa rhomboidea, которая является дном четвертого желудочка.
Мозжечок: внешнее строение и функции отдела ствола головного мозга человека
Мозжечок (малый мозг), cerebellum, располагается кзади (дорсальнее) от моста и от верхней дорсальной части продолговатого мозга. Мозжечок ствола головного мозга лежит в задней черепной ямке.
Внешнее строение мозжечка головного мозга:
- Полушария, hemispheric! cerebelli;
- Горизонтальная борозда, sulcus horizontalis, разделяет верхнюю и нижнюю поверхности полушарий;
- Червь, vermis: верхний червь, vermis superior, и нижний червь, vermis inferior;
- Пластиночки, lamellae (извилины, gyri), объединяются в пластины, laminae, которые получили название — листки мозжечка, folia cerebelli; последние группируются в дольки, lobuli.
Поверхность мозжечка головного мозга имеет своеобразный рельеф. По ней проходят параллельно расположенные щели, подразделяя ее на листки, которые группируются в дольки мозжечка. Каждой дольке червя соответствуют определенные дольки полушарий. Поверхность полушарий покрыта корой. В белом веществе заложены ядра: ядро шатра, шаровидное, пробковидное, зубчатое ядра. Каждое ядро связано с определенной зоной коры мозжечка.
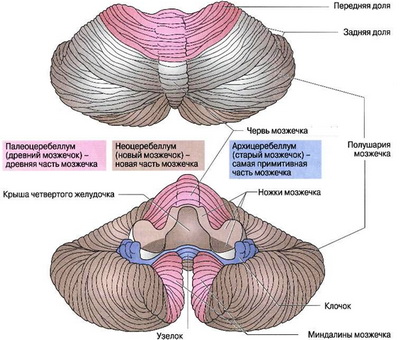
С филогенетической точки зрения мозжечок головного мозга человека делится на 3 части. Старый мозжечок, archeocerebellum, представлен узелком в черве и клочком в полушариях; вместе они составляют клочково-узелковуюдолю, lobusflocculonodularis. Эта часть мозжечка связана с преддверными ядрами и ретикулярной формацией и имеет отношение к поддержанию равновесия и пространственной ориентации тела.
Древний мозжечок, paleocerebellum, включает нижнюю часть червя и прилегающие к нему участки полушарий. Этот отдел мозжечка получает проприоцептивные сигналы из спинного мозга, связан с оливами и различными ядрами ствола головного мозга. Функция его заключается, в основном, в участии в регуляции мышечного тонуса.
Новый мозжечок, neocerebellum, составляют верхняя часть червя и большая часть полушарий. Он связан с корой большого мозга через ядра моста. Его функциональное значение состоит в контроле и координации произвольных движений, прежде всего моторики конечностей.
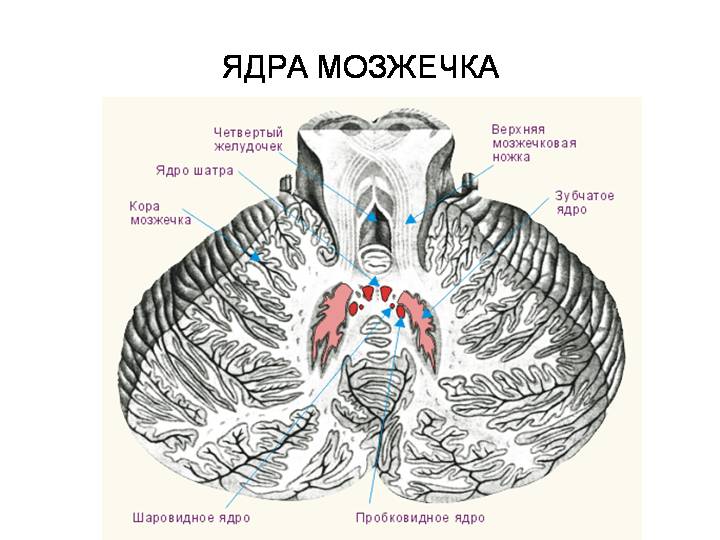 Мозжечок соединяется с отделом ствола головного мозга посредством 3 пар ножек. Нижние мозжечковые ножки идут от продолговатого мозга, они содержат в основном афферентные пути: задний спинно-мозжечковый, преддверно-мозжечковый и оливомозжечковый. Эти пути заканчиваются в старом и древнем мозжечке. В нижних ножках проходят также эфферентный мозжечково-ядерный путь от ядра шатра к латеральному пред-дверному ядру, двигательным ядрам черепных нервов, ядрам ретикулярной формации. Средние мозжечковые ножки состоят из мосто-мозжечковых волокон. Они оканчиваются в коре нового мозжечка. В составе верхних мозжечковых ножек преобладают эфферентные пути, направляющиеся от зубчатого ядра мозжечка к красному ядру среднего мозга (зубчато-красноядерный) и ядрам таламуса (зубчато-таламический). К афферентным путям относится передний спинно-мозжечковый путь, оканчивающийся в древнем мозжечке. В верхних ножках содержатся также волокна, соединяющие непосредственно кору мозжечка и большого мозга.
Мозжечок соединяется с отделом ствола головного мозга посредством 3 пар ножек. Нижние мозжечковые ножки идут от продолговатого мозга, они содержат в основном афферентные пути: задний спинно-мозжечковый, преддверно-мозжечковый и оливомозжечковый. Эти пути заканчиваются в старом и древнем мозжечке. В нижних ножках проходят также эфферентный мозжечково-ядерный путь от ядра шатра к латеральному пред-дверному ядру, двигательным ядрам черепных нервов, ядрам ретикулярной формации. Средние мозжечковые ножки состоят из мосто-мозжечковых волокон. Они оканчиваются в коре нового мозжечка. В составе верхних мозжечковых ножек преобладают эфферентные пути, направляющиеся от зубчатого ядра мозжечка к красному ядру среднего мозга (зубчато-красноядерный) и ядрам таламуса (зубчато-таламический). К афферентным путям относится передний спинно-мозжечковый путь, оканчивающийся в древнем мозжечке. В верхних ножках содержатся также волокна, соединяющие непосредственно кору мозжечка и большого мозга.
Самые древние связи мозжечка установились с органами равновесия. От вестибулярных ядер протянулись нервные волокна, представляющие собой часть преддверно-мозжечкового пути. Они проникают в мозжечок в составе его нижних ножек и заканчиваются на нейронах коры клочка и узелка. От нейронов этих участков коры мозжечка начинается нисходящий (эфферентный) путь. Нервные волокна из коры клочка и узелка достигают нейронов ядра шатра, которое является древнейшим из ядер мозжечка. Аксоны нейронов ядра шатра через нижние ножки мозжечка достигают ядер ретикулярной формации продолговатого мозга. От них по ретикулярно-спинйомозговому пути эфферентные импульсы поступают к мышцам туловища. Описанные нервные связи мозжечка (преддверно-мозжечковый афферентный путь и мозжечково-ретикулярно-спинномозговой путь) играли важную роль у обитателей водной среды, локомоция которых осуществлялась главным образом за счет мышц туловища.
Позже, в связи с выходом животных на сушу и развитием конечностей, появились афферентные пути от рецепторов мышц, сухожилий и суставов конечностей в виде заднего и переднего спинно-мозжечковых путей. Волокна заднего спинно-мозжечкового пути (пучок Флексига) проходят в мозжечок в составе нижних ножек мозжечка и заканчиваются на нейронах коры нижней части червя. Волокна переднего спинно-мозжечкового пути (пучок Говерса) входят в мозжечок в составе его верхних ножек и заканчиваются на нейронах коры верхней части червя. Эфферентный путь от нейронов коры червя идет к нейронам шаровидного и пробковидного ядер мозжечка. Аксоны нейронов этих ядер выходят из мозжечка в составе его нижних ножек и достигают нейронов ретикулярной формации. В дальнейшем эфферентные пути направляются к мышцам конечностей.
В связи с усложнением функций мускулатуры туловища и конечностей также сформировались бульбарно-мозжечковые и ядерно-мозжечковые пути, которые заканчиваются в коре средней части червя мозжечка. Эфферентный путь от нейронов средней части коры червя аналогичен предыдущим.
У млекопитающих, и особенно у человека, получила развитие система так называемых предмозжечковых ядер: ядра оливы и собственные ядра моста. Ядра оливы получают импульсы по коллатералям, отходящим от волокон пирамидных и экстра-пирамидных путей. Аксоны нейронов оливы проходят через нижние ножки мозжечка, формируя оливо-мозжечковый путь. Волокна этого пути переходят в продолговатом мозге на противоположную сторону и заканчиваются на нейронах коры полушарий мозжечка.
 Собственные ядра моста являются коммуникационными центрами по ходу корково-мосто-мозжечкового пути. На нейронах этих ядер заканчиваются волокна корково-мостовых путей, а также коллатерали пирамидных волокон. Аксоны нейронов собственных ядер моста переходят на противоположную сторону и в составе средних ножек мозжечка направляются к нейронам коры полушарий мозжечка. Из коры полушарий мозжечка начинается мозжечково-зубчато-красноядерно-спинномозговой путь, который осуществляет так называемую «поправочную» деятельность при выполнении сложных произвольных движений, главным образом, верхней конечностью. Сложные нервные связи мозжечка с корой полушарий большого мозга объясняют разнообразие его функций. Это не просто «орган равновесия».
Собственные ядра моста являются коммуникационными центрами по ходу корково-мосто-мозжечкового пути. На нейронах этих ядер заканчиваются волокна корково-мостовых путей, а также коллатерали пирамидных волокон. Аксоны нейронов собственных ядер моста переходят на противоположную сторону и в составе средних ножек мозжечка направляются к нейронам коры полушарий мозжечка. Из коры полушарий мозжечка начинается мозжечково-зубчато-красноядерно-спинномозговой путь, который осуществляет так называемую «поправочную» деятельность при выполнении сложных произвольных движений, главным образом, верхней конечностью. Сложные нервные связи мозжечка с корой полушарий большого мозга объясняют разнообразие его функций. Это не просто «орган равновесия».
Необходимо отметить, что эфферентные пути мозжечка перекрещиваются два раза (до красного ядра и после красного ядра). В результате этого одностороннее поражение мозжечка проявляется выпадением функции мышц одноименной стороны. Даже односторонние поражения мозжечка вызывают нарушения равновесия тела.
Мозжечок (cerebellum, малый мозг) — одна из интегративных структур головного мозга, основная функция мозжечка – участие в координации и регуляции произвольных, непроизвольных движений, в регуляции вегетативных и поведенческих функций.
Приходящая в мозжечок афферентная сигнализация от статорецепторов внутреннего уха, проприорецепторов скелетной мускулатуры подвергается автоматической обработке и сопоставляется с импульсами, посылаемыми корой большого мозга. Выполняя свои функции, мозжечок головного мозга человека учитывает влияние силы тяжести и инерции при движениях тела и вносит соответствующие поправки в двигательные акты. Это определяет роль мозжечка в координации движений. При поражениях мозжечка наблюдаются нарушения согласованных движений (асинергия), несоразмерность движений (дизметрия), снижение тонуса мышц, шаткость походки (атаксия) и другие двигательные расстройства.
IV желудочек полости ромбовидного мозга
IV желудочек, ventriculus quartus, является полостью ромбовидного мозга. Он представляет собой продолжение центрального канала спинного мозга.
Крыша IV желудочка имеет две части, различающиеся по развитию и строению. В области продолговатого мозга дорсальная стенка нервной трубки атрофируется и остается в виде эпендимального эпителия. К наружной поверхности эпителиальной пластинки, lamina epitelialis, прилегает дупликатура сосудистой оболочки IV желудочка, tela choroidea ventriculi quarti.
Нижний (задний) мозговой парус, velum medullare inferius, ромбовидного мозга — парный, натянут между узелком червя, ножкой клочка и клочком.
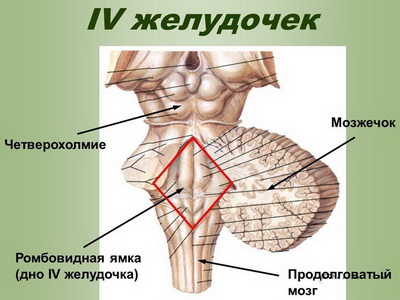 В области моста из дорсальной стенки нервной трубки развивается мозжечок. Передняя часть крыши IV желудочка образована пластинкой белого вещества — верхним (передним) мозговым парусом, velum medullare superius, который натянут между верхними ножками мозжечка.
В области моста из дорсальной стенки нервной трубки развивается мозжечок. Передняя часть крыши IV желудочка образована пластинкой белого вещества — верхним (передним) мозговым парусом, velum medullare superius, который натянут между верхними ножками мозжечка.
Таким образом, задняя часть крыши IV желудочка представлена задним мозговым парусом и мягкой мозговой оболочкой. Последняя сращена со свободным краем заднего мозгового паруса, с нижними ножками мозжечка и задними канатиками спинного мозга.
Полость IV желудочка внизу сообщается с центральным каналом спинного мозга, вверху — с водопроводом среднего мозга. Кроме того, в области латеральных углов ромбовидной ямки в сосудистой основе IV желудочка имеется сообщение с подпаутинным межоболочечным пространством через парное отверстие, которое называется боковой апертурой, apertura lateralis (отверстие Люшка). Еще одно непарное отверстие имеется в области нижнего угла ромбовидной ямки — срединная апертура, apertura mediana (отверстие Мажанди). Через указанные отверстия спинномозговая жидкость поступает из четвертого желудочка в подпаутинное межоболочечное пространство головного мозга.
Дно IV желудочка мозга: ромбовидная ямка
Ромбовидная ямка, rhomboidea, образует дно IV желудочка, имеет, действительно, форму ромба, ограниченного верхними и нижними ножками мозжечка. В ней выделяют две половины: нижнюю (каудальную) и верхнюю (ростральную), разграниченные мозговыми полосками, striae medullares. Нижняя половина ромбовидной ямки мозга является дорсальной поверхностью продолговатого мозга, верхняя — дорсальной поверхностью моста.
По срединной линии ромбовидной ямки проходит срединная борозда, sulcus medianus, по обе стороны от которой имеется продольной формы медиальное возвышение, eminentia medialis. Латерально оно ограничено пограничной бороздой, sulcus limitans. Эта борозда имеет важное значение, так как служит примерной границей между проекцией двигательных и чувствительных ядер черепных нервов. Двигательные ядра проецируются медиальнее, чувствительные ядра — латеральнее. Медиальное возвышение в нижнем углу ромбовидной ямки называют треугольником подъязычного нерва, trigonum nervi hypoglossi. Выше мозговых полосок медиальное возвышение образует заметное утолщение, называемое лицевым бугорком, coliiculusfacialis.
В пределах ромбовидной ямки также проецируются ядра ретикулярной формации, в частности, в верхнелатеральной ее части выделяют голубое пятно, locus coeruleus, по срединной линии продолговатого мозга — ядра срединного шва, nuclei raphe mediani.
Закономерности топографии ядер и проекции их на ромбовидную ямку
 Ближе к срединной линии располагаются двигательные ядра, латеральнее — вегетативные и еще дальше — чувствительные ядра, что связано с раскрытием краниального конца нервной трубки в эмбриогенезе, в результате чего ядра, соответствующие задним столбам спинного мозга, смещаются кнаружи. Ядра V— VIII пар черепных нервов заложены преимущественно в мосте и проецируются на верхний треугольник ромбовидной ямки. Ядра IX—XII парчерепных нервов располагаются преимущественно в продолговатом мозге и проецируются на нижний треугольник ромбовидной ямки.
Ближе к срединной линии располагаются двигательные ядра, латеральнее — вегетативные и еще дальше — чувствительные ядра, что связано с раскрытием краниального конца нервной трубки в эмбриогенезе, в результате чего ядра, соответствующие задним столбам спинного мозга, смещаются кнаружи. Ядра V— VIII пар черепных нервов заложены преимущественно в мосте и проецируются на верхний треугольник ромбовидной ямки. Ядра IX—XII парчерепных нервов располагаются преимущественно в продолговатом мозге и проецируются на нижний треугольник ромбовидной ямки.
Таким образом, чувствительные ядра ромбовидного мозга, соответствующие задним рогам спинного мозга, занимают в ромбовидной ямке латеральное положение. Двигательные ядра, соответствующие передним рогам спинного мозга, располагаются в ромбовидной ямке медиально. В белом веществе между двигательными и чувствительными ядрами ромбовидной ямки находятся ядра автономной (вегетативной) нервной системы.
Следующий раздел статьи посвящен строению и функциям среднего мозга.
Средний головной мозг человека: строение поверхностей и функции отдела
Верхней (передней) границей среднего головного мозга на его вентральной поверхности служат зрительные тракты и сосцевидные тела, на задней — передний край моста. На дорсальной поверхности верхняя (передняя) граница среднего мозга соответствует задним краям (поверхностям) таламусов, задняя (нижняя) — уровню выхода корешков блокового нерва (nervus trochlearis, IV пара).
Средний мозг человека подразделяется на крышу и ножки большого мозга. Крыша среднего мозга имеет верхние и нижние холмики. Полостью среднего мозга является водопровод мозга (Сильвиев водопровод).
Образования на вентральной поверхности в строении среднего мозга:
- Ножки мозга, pedunculi cerebri;
- Мжножковая ямка, fossa interpeduncularis;
- Борозда глазодвигательного нерва, sulcus oculomotorius;
- Заднее продырявленное вещество, substantia perforata posterior, (расположено в межножковой ямке,/гшд interpeduncularis).
Образования на дорсальной поверхности в отделе среднего мозга:
- Пластинка крыши среднего мозга, lamina tecti:
- верхние холмики, colliculisuperiores’,
- нижние холмики, colliculi inferiores;
- ручки верхних холмиков, brachia colliculisuperiores, (связаны с латеральными коленчатыми телами);
- ручки нижних холмиков, brachia colliculi inferiores, (связаны с медиальными коленчатыми телами);
- Уздечка верхнего мозгового паруса, frenulum velli medullaris superioris.
Функция среднего мозга заключается в повышении тонуса мышц. Активация среднего мозга через гипоталамус приводит к учащению сокращений сердца; происходит подготовка к избеганию, к оборонительной реакции.
Четверохолмие организует ориентировочные зрительные и слуховые рефлексы. У человека четверохолмный рефлекс является сторожевым. В случаях повышенной возбудимости четверохолмий при внезапном звуковом или световом раздражении у человека возникает вздрагивание, иногда вскакивание на ноги, вскрикивание, максимально быстрое удаление от раздражителя, подчас безудержное бегство.
При нарушении четверохолмного рефлекса человек не может быстро переключаться с одного вида движения на другое. Следовательно, четверохолмия принимают участие в организации произвольных движений.
Красные ядра связаны с корой большого мозга (нисходящие от коры пути), подкорковыми ядрами, мозжечком, спинным мозгом (красноядерно-спинномозговой путь).
Базальные ганглии головного мозга, мозжечок имеют свои окончания в красных ядрах.
Структура среднего головного мозга: ядра черепных нервов
Собственно ножка мозга отделяется от покрышки черным веществом, относящимся к ядрам экстрапирамидной системы. В ножке мозга проходят нисходящие пути от коры. Медиальный и латеральный участки ножки мозга занимают корково-мостовые волокна, а в средней части проходит пирамидный путь. Дорсальная часть содержит в основном филогенетически более старые образования.
В покрышке ножки среднего мозга глазодвигательный нерв имеет три ядра. Двигательное ядро, nucleus nervus oculomotorii, — наиболее крупное, имеет вытянутую форму, парное.
 Кроме парного двигательного ядра у глазодвигательного нерва среднего отдела головного мозга имеется еще одно непарное ядро — центральное непарное ядро, nucleus centralis impar. Данное ядро взаимосвязано с каудальными сегментами основных двигательных ядер обеих сторон, отвечающих за иннервацию медиальных прямых мышц. При этом обеспечивается сочетанная работа указанных мышц правого и левого глазных яблок, которые вращают глазное яблоко и приближают зрачки к срединной плоскости. В связи со своей функцией центральное непарное ядро называют также конвергенционным.
Кроме парного двигательного ядра у глазодвигательного нерва среднего отдела головного мозга имеется еще одно непарное ядро — центральное непарное ядро, nucleus centralis impar. Данное ядро взаимосвязано с каудальными сегментами основных двигательных ядер обеих сторон, отвечающих за иннервацию медиальных прямых мышц. При этом обеспечивается сочетанная работа указанных мышц правого и левого глазных яблок, которые вращают глазное яблоко и приближают зрачки к срединной плоскости. В связи со своей функцией центральное непарное ядро называют также конвергенционным.
Дорсальнее от двигательных ядер вблизи срединной линии располагается парное вегетативное ядро глазодвигательного нерва — так называемое добавочное ядро, nucleus oculomotorius accessorius (ядро Якубовича). Нейроны этого ядра отвечают за иннервацию мышцы, суживающей зрачок, и ресничной мышцы.
Часть волокон от двигательных соматических ядер глазодвигательного нерва участвует в образовании медиального продольного пучка. Большая часть волокон от всех ядер составляет корешок глазодвигательного нерва, который выходит из вещества мозга в одноименной борозде.
Между центральным серым веществом и красными ядрами располагается ретикулярная формация, содержащая многочисленные мелкие ядра и два крупных ядра. Одно из них называется промежуточным ядром, nucleus interstitialis (ядро Кахаля), второе — ядром задней спайки мозга, nucleus commissurae posterior (ядро Даркшевича). Аксоны клеток ядра Кахаля и ядра Даркшевича направляются в спинной мозг, формируя при этом медиальный продольный пучок, fasciculus longitudinalis medialis.
В составе медиального продольного пучка проходят нервные волокна, обеспечивающие связь ядер ретикулярной формации и двигательных ядер III, IV, VI и XI пар черепных нервов среднего мозга. Следовательно, ядро Кахаля и ядро Даркшевича среднего мозга являются центрами координации сочетанной функции мышц глазного яблока и мышц шеи. Так как функция этих мышц в наибольшей степени проявляется при вестибулярных нагрузках, к ядрам ретикулярной формации поступают афферентные импульсы от вестибулярных ядер моста (ядра VIII пары черепных нервов).
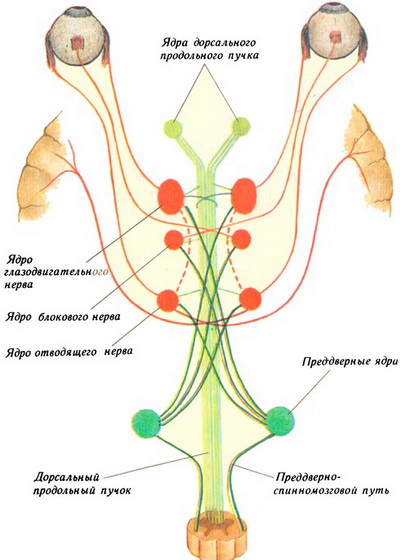 Рядом с медиальным продольным пучком располагается дорсальный продольный пучок, fasciculus longitudinalis dorsalis, который начинается от структур среднего промежуточного мозга. Волокна этого пучка направляются к вегетативным ядрам черепных нервов и спинного мозга. Они обеспечивают координацию деятельности вегетативных центров ствола головного и спинного мозга.
Рядом с медиальным продольным пучком располагается дорсальный продольный пучок, fasciculus longitudinalis dorsalis, который начинается от структур среднего промежуточного мозга. Волокна этого пучка направляются к вегетативным ядрам черепных нервов и спинного мозга. Они обеспечивают координацию деятельности вегетативных центров ствола головного и спинного мозга.
Дорсальнее водопровода мозга располагается крыша среднего мозга. Ее составляют две пары холмиков — верхние и нижние, colliculi superiores et inferiores, которые существенно различаются по строению. У человека более развиты верхние холмики, так как основную часть информации он получает через посредство органа зрения. Верхние холмики представляют собой интеграционный центр среднего мозга и, кроме того, являются одним из подкорковых центров зрения, обоняния и тактильной чувствительности.
На нейронах ядер нижних холмиков заканчиваются три четверти волокон латеральной петли. Они являются подкорковыми центрами слуха. Часть волокон латеральной петли в составе ручек нижних холмиков направляется в ядро медиального коленчатого тела промежуточного мозга. Еще некоторое количество волокон латеральной петли заканчивается в верхних холмиках. Туда же поступает и часть волокон обонятельного пути, а также часть афферентных волокон, идущих в составе lemniscus spinalis et lemniscus medialis. Верхние холмики имеют выраженную слоистость расположения нейронов, что характерно для интеграционных центров ствола головного мозга (кора мозжечка и кора полушарий большого мозга).
В поверхностных слоях верхних холмиков заканчиваются волокна зрительных трактов. В глубоких слоях происходит последовательное синаптическое переключение волокон.
Аксоны нейронов глубоких слоев в структуре среднего головного мозга образуют пучок, который располагается латеральнее центрального серого вещества. В составе пучка проходят два тракта — покрышечно-спинномозговой путь, tractus tectospinalis, и покрышечно-ядерный пучок, fasciculus tectonuclearis. Волокна этих путей переходят на противоположную сторону, образуя, дорсальный перекрест покрышки, decussatio tegmenti dorsalis (перекрест Мейнерта), который находится вентрально по отношению к Сильвиеву водопроводу и имеет еще одно название — фонтановидный.
Волокна покрышечно-спинномозгового пути заканчиваются на нейронах собственных ядер передних рогов спинного мозга. Волокна покрышечно-ядерного пучка заканчиваются на нейронах двигательных ядер черепных нервов. Покрышечноспинномозговой и покрышечно-ядерный пути проводят нервные импульсы, обеспечивающие выполнение защитных рефлекторных движений (настораживание, вздрагивание, прыжок в сторону) в ответ на различные сильные раздражения (зрительные, слуховые, обонятельные и тактильные).
